Главная → Физиология → Механизм регуряции физиологических процессов -> Функции ретикулярной формации
Содержание
Функции ретикулярной формации
К собственно интегративных функций ретикулярной формации относятся:
- контроль над состояниями сна и бодрствования
- мышечный (фазный и тонический) контроль
- обработка информационных сигналов окружающей и внутренней среды организма, которые поступают по разным каналам
Ретикулярная формация объединяет различные участки ствола мозга (ретикулярную формацию продолговатого мозга, варолиева моста и среднего мозга). В функциональном отношении в ретикулярной формации разных отделов мозга есть много общего, поэтому целесообразно рассматривать ее как единую структуру. Ретикулярная формация представляет собой диффузное накопление клеток разного вида и величины, которые разделены многими волокнами. Кроме этого, в середине ретикулярной формации выделяют около 40 ядер и пидьядер. Нейроны ретикулярной формации имеют широко разветвленные дендриты и продолговатые аксоны, часть которых делится Т-образно (один отросток направлен вниз, образуя ретикулярный-спинальный путь, а второй — в верхние отделы головного мозга).
В ретикулярной формации сходится большое количество афферентных путей из других мозговых структур: из коры большого мозга — коллатерали кортико-спинальных (пирамидных) путей, из мозжечка и других структур, а также коллатеральные волокна, которые подходят через ствол мозга, волокна сенсорных систем (зрительные, слуховые и т.д.). Все они заканчиваются синапсами на нейронах ретикулярной формации. Так, благодаря такой организации ретикулярная формация приспособлена к объединению влияний из различных структур мозга и способна влиять на них, то есть выполнять интегративные функции в деятельности ЦНС, определяя в значительной мере общий уровень ее активности.
Свойства ретикулярных нейронов. Нейроны ретикулярной формации способны к устойчивой фоновой импульсной активности. Большинство из них постоянно генерирует разряды частотой 5-10 Гц. Причиной такой постоянной фоновой активности ретикулярных нейронов являются: во-первых, массивная конвергенция различных афферентных влияний (от рецепторов кожных, мышечных, висцеральных, глаза, уши и др.)., А также воздействий из мозжечка, коры большого мозга, вестибулярных ядер и других мозговых структур на один и тот же ретикулярный нейрон. При этом зачастую в ответ на это возникает возбуждение. Во-вторых, активность ретикулярного нейрона может быть изменена гуморальными факторами (адреналин, ацетилхолин, напряжение С02 в крови, гипоксия и др.).. Эти непрерывные импульсы и химические вещества, содержащиеся в крови, поддерживают деполяризацию мембран ретикулярных нейронов, их способность к устойчивой импульсной активности. В связи с этим ретикулярная формация тоже оказывает на другие мозговые структуры постоянный тонический влияние.
Характерной особенностью ретикулярной формации также высокая чувствительность ее нейронов в различных физиологически активных веществ. Благодаря этому деятельность ретикулярных нейронов может быть сравнительно легко блокирована фармакологическими препаратами, которые связываются с циторецепторамы мембран этих нейронов. Особенно активными в этом отношении соединения барбитуровой кислоты (барбитураты), аминазин и другие лекарственные препараты, которые широко применяются в медицинской практике.
Характер неспецифических влияний ретикулярной формации. Ретикулярная формация ствола мозга участвует в регуляции вегетативных функций организма. Однако еще в 1946 г. американский нейрофизиолог Н. W. Megoun и его сотрудники обнаружили, что ретикулярная формация имеет непосредственное отношение к регуляции соматической рефлекторной деятельности. Было доказано, что ретикулярная формация оказывает диффузное неспецифическое, нисходящее и восходящее влияние на другие мозговые структуры.
Нисходящее влияние. При раздражении ретикулярной формации заднего мозга (особенно гигантоклеточной ядра продолговатого мозга и ретикулярного ядра моста, где принимают начало ретикулоспинальному пути), возникает торможение всех спинальных двигательных центров (сгибательных и разгибательных). Это торможение очень глубокое и продолжительное. Такое положение в естественных условиях может наблюдаться при глубоком сне.
Наряду с диффузными тормозящими влияниями, при раздражении определенных участков ретикулярной формации выявляется диффузное влияние, которое облегчает деятельность спинальной двигательной системы.
Ретикулярная формация играет важную роль в регуляции деятельности мышечных веретен, изменяя частоту разрядов, поступающие гамма-эфферентными волокнами к мышцам. Таким образом модулируется обратная импульсация в них.
Восходящий влияние. Исследования Н. W. Megoun, G. Moruzzi (1949) показали, что раздражение ретикулярной формации (заднего, среднего и промежуточного мозга) сказывается на деятельности высших отделов головного мозга, в частности коры большого мозга, обеспечивая переход ее в активное состояние. Это положение подтверждается данными многочисленными экспериментальными исследованиями и клиническими наблюдениями. Так, если животное находится в состоянии сна, то прямое раздражение ретикулярной формации (особенно варолиева моста) через введенные в эти структуры электроды вызывает поведенческую реакцию пробуждения животного. При этом на ЭЭГ возникает характерное изображение — изменение альфа-ритма бета-ритмом, т.е. фиксируется реакция десинхронизации или активизации. Указанная реакция не ограничивается определенным участком коры большого мозга, а охватывает большие ее массивы, т.е. носит генерализованный характер. При разрушении ретикулярной формации или выключении ее восходящих связей с корой большого мозга животное впадает в сноподобное состояние, не реагирует на световые и обонятельные раздражители, фактически не вступает в контакт с внешним миром. То есть конечный мозг прекращает активно функционировать.
Таким образом, ретикулярная формация ствола головного мозга выполняет функции восходящей активирующей системы мозга, которая поддерживает на высоком уровне возбудимость нейронов коры большого мозга.
Кроме ретикулярной формации ствола мозга, в восходящую активирующую систему головного мозга входят также неспецифические ядра таламуса, задний гипоталамус, лимбических структуры. Являясь важным интегративным центром, ретикулярная формация, в свою очередь, является частью более глобальных интеграционных систем мозга, которые включают гипоталамо-лимбические и неокортикальные структуры. Именно во взаимодействии с ними и формируется целесообразное поведение, направленное на приспособление организма к меняющимся условиям внешней и внутренней среды.
Одним из основных проявлений повреждения ретикулярных структур у человека является потеря сознания. Она бывает при черепно-мозговых травмах, нарушении мозгового кровообращения, опухолях и инфекционных процессах в стволе мозга. Длительность состояния обморока зависит от характера и выраженности нарушений функции ретикулярной активизирующей системы и колеблется от нескольких секунд до многих месяцев. Дисфункция восходящих ретикулярных влияний проявляется тоже потерей бодрости, постоянной патологической сонливостью или частыми приступами засыпания (пароксизмальная гиперсомия), беспокойным ночным сном. Наблюдаются также нарушения (чаще повышении) мышечного тонуса, различные вегетативные изменения, эмоционально-психические расстройства и др.
Ретикулярная формация ствола головного мозга
Ретикулярная формация (лат. rete — сеть) представляет собой совокупность клеток, клеточных скоплений и нервных волокон, расположенных на всем протяжении ствола мозга (продолговатый мозг, мост, средний и промежуточный мозг) и в центральных отделах спинного мозга . Ретикулярная формация получает информацию от всех органов чувств , внутренних и других органов , оценивает ее, фильтрует и передает в лимбическую систему и кору большого мозга.
Ретикулярная формация: особенности и функции
Она регулирует уровень возбудимости и тонуса различных отделов центральной нервной системы, включая кору большого мозга, играет важную роль в сознании, мышлении, памяти, восприятии, эмоциях, сне, бодрствовании, вегетативных функциях, целенаправленных движениях, а также в механизмах формирования целостных реакций организма. Ретикулярная формация прежде всего выполняет функцию фильтра, который позволяет важным для организма сенсорным сигналам активировать кору мозга, но не пропускает привычные для него или повторяющиеся сигналы.
Ретикулярная формация представляет собой важный пункт на пути восходящей неспецифической соматосенсорной системы . Соматовисцеральные афференты идут в составе спиноретикулярного тракта ( переднебоковой канатик ), а также, возможно, в составе проприоспинальных (полисинаптических) путей и соответствующих путей от ядра спинального тройничного тракта . К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов , т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделов головного мозга — от моторных областей коры и сенсорных областей коры , от таламуса и гипоталамуса . Имеется также множество эфферентных связей — нисходящие к спинному мозгу и восходящие через неспецифические таламические ядра к коре головного мозга , гипоталамусу и лимбической системе . Большинство нейронов образует синапсы с двумя — тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации. Другими их свойствами являются большие рецептивные поля поверхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции (стохастические колебания числа потенциалов действия при повторной стимуляции). Все эти свойства противоположны свойствам лемнисковых нейронов в специфических ядрах соматосенсорной системы ( рис.9-7 и рис. 5-13 ).
Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
1. в регуляции уровня сознания путем воздействия на активность корковых нейронов , например, участие в цикле сон / бодрствование ,
2. в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам , идущим по переднебоковому канатику , путем проведения афферентной информации к лимбической системе ,
3. в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах ( циркуляторных рефлексах и дыхательных рефлексах , рефлекторных актах глотания , кашля , чихания ), при которых должны взаимно координироваться разные афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга .
Ссылки:
Все ссылки
Ретикулярная формация известна с 1845 г., описана Дейтерсом (O.F.C. Deiters) в 1885 г. В настоящее время продолжается ее изучение. Ретикулярная формация располагается между задними и боковыми рогами шейных сегментов спинного мозга, в покрышке мозгового ствола, в центральном ядре зрительного бугра. Представляет собой комплекс анатомически и функционально взаимосвязанных нейронов, окруженных множеством волокон, идущих в различных направлениях к ядерным структурам и проводящим путям (рис.
Ретикулярная формация
30).
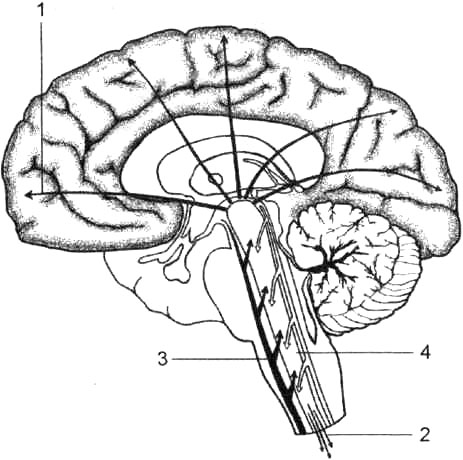 Рис. 30. Ретикулярная формация ствола мозга: 1 — восходящие пути; 2 — нисходящие пути; 3 — специфический (лемнисковый) чувствительный проводящий путь; 4 — пирамидальный путь.
Рис. 30. Ретикулярная формация ствола мозга: 1 — восходящие пути; 2 — нисходящие пути; 3 — специфический (лемнисковый) чувствительный проводящий путь; 4 — пирамидальный путь.
Ретикулярная формация воспринимает все импульсы (болевые, температурные, световые, звуковые и пр.), однако в ней нет специализированных нейронов. Поэтому одни и те же нейроны воспринимают различные импульсы и передают их в различные отделы мозга, во все участки коры. Ретикулярная формация — это вторая афферентная система головного мозга, его неспецифическая структура. Она имеет двухсторонние связи со всеми структурами головного и спинного мозга (рис. 31, 32).
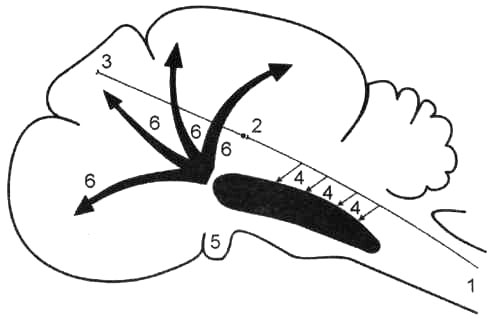 Рис. 31. Схема восходящего активирующего влияния ретикулярной формации: 1; 2; 3 — специфический (лемнисковый) чувствительный проводящий путь; 4 — коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 — восходящая активирующая система ретикулярной формации; 6 — генерализованное влияние ретикулярной формации на кору головного мозга.
Рис. 31. Схема восходящего активирующего влияния ретикулярной формации: 1; 2; 3 — специфический (лемнисковый) чувствительный проводящий путь; 4 — коллатерали, соединяющие специфический чувствительный путь с ретикулярной формацией мозгового ствола; 5 — восходящая активирующая система ретикулярной формации; 6 — генерализованное влияние ретикулярной формации на кору головного мозга.
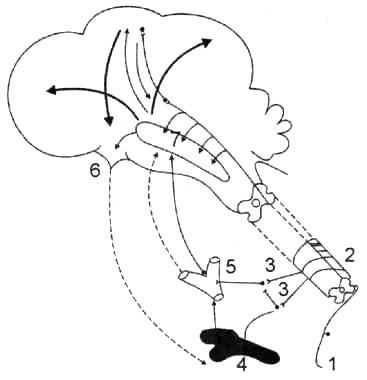 Рис.
Рис.
32. Схема взаимодействия между симпатическим тонусом и электрической активностью коры головного мозга: 1 — чувствительный нерв, на который наносится стимул (болевое раздражение); 2 — спинной мозг; 3 — симпатические нервы; 4 — надпочечник; 5 — каротидный синус; 6 — гипофиз; 7 — ретикулярная формация. Сплошными стрелками обозначены нервные влияния, пунктирными — гормональные влияния, которые через ретикулярную формацию оказывают активирующее воздействие на кору головного мозга.
Структурные элементы ретикулярной формации ствола мозга подразделяют на латеральный и медиальный отделы. В латеральном отделе заканчиваются волокна из различных афферентных систем. К рассеянным клеткам и ядрам ретикулярной формации подходят коллатерали от медиальной и латеральной петель, от чувствительных ядер черепных нервов. От нейронов медиального отдела начинаются эфферентные волокна к двигательным ядрам черепных нервов, к мозжечку, к двигательным ядрам передних рогов спинного мозга.
Основные афферентные пути ретикулярной формации: tr. spinoreticularis — от спинного мозга, tr. tegmentothalamicus — от среднего мозга, reticulothalamicus — от продолговатого мозга и моста, tr. thalamocorticalis — ко всем областям и слоям коры полушарий большого мозга. Сетевидная формация активизирует кору большого мозга и мозжечок.
Кора полушарий большого мозга, в свою очередь, посылает по tr. corticoreticularis импульсы в ретикулярную формацию в составе пирамидных путей. Основным эфферентным трактом является tr. reticulospinalis. Этот путь проводит тонические импульсы к гамма-мотонейронам спинного мозга. Ретикулярная формация регулирует моторное звено, обеспечивая координацию движений, синхронность мышечных сокращений, обеспечивает нестандартные движения, рефлекс равновесия, устанавливает антигравитационный тонус мускулатуры, удерживающий тело над землей. Ретикулярная формация перераспределяет мышечный тонус, что в кризисных ситуациях приводит к мобилизации скрытых резервов организма.
Установлена роль голубоватого пятна и ядер шва в регуляции сна и бодрствования. Голубоватое пятно (locus caeruleus) находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют норадреналин, активирующий вышележащие отделы мозга. Особенно высока активность нейронов голубоватого пятна во время бодрствования, во время глубокого сна она угасает почти полностью.
Ядра шва (nuclei raphes) располагаются по срединной линии продолговатого мозга. Нейроциты этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Ядра ретикулярной формации продолговатого мозга имеют связи с вегетативными ядрами Ⅸ, Ⅹ нервов и симпатическими ядрами спинного мозга. Поэтому они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и так далее.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, к медиальному продольному пучку (fasciculus longitudinalis medialis), имеют связи с ядрами третьей, четвертой, шестой, восьмой, девятой, десятой и одиннадцатой парой черепных нервов. Они координируют работу этого пучка, обеспечивая сочетанные повороты головы и глаз при изменении позы или при поиске источника звука, фиксацию взгляда. (Данные движения совершенно необходимы при трудовых и игровых актах).
Эти связи объясняют вегетативные нарушения при вестибулярных перегрузках. Рассеянные нейроны ретикулярной формации выступают в качестве вставочных нейронов охранительных рефлексов глотательного, роговичного (рис. 33), кашлевого рвоты, зевоты, чихания и др.
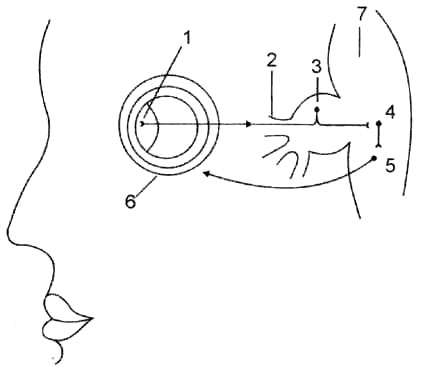 Рис. 33. Схема роговичного рефлекса: 1 — рецепторы, расположенные в роговице; 2 — глазная ветвь тройничного нерва; 3 — ложноуниполярная клетка чувствительного узла тройничного нерва; 4 — ассоциативный нейрон — рассеянная клетка ретикулярной формации; 5 — клетка двигательного ядра лицевого нерва; 6 — круговая мышца глаза.
Рис. 33. Схема роговичного рефлекса: 1 — рецепторы, расположенные в роговице; 2 — глазная ветвь тройничного нерва; 3 — ложноуниполярная клетка чувствительного узла тройничного нерва; 4 — ассоциативный нейрон — рассеянная клетка ретикулярной формации; 5 — клетка двигательного ядра лицевого нерва; 6 — круговая мышца глаза.
Тема 13. Ретикулярная формация.
Термин ретикулярная формация предложил в 1865 году немецкий ученый О. Дейтерс. Под этим термином Дейтерс понимал разбросанные в стволе головного мозга клетки, окруженные множеством волокон, идущих в различных направлениях. Именно сетевидное расположение волокон, связывающих между собой нервные клетки, послужило основой для предложенного названия.
В настоящее время морфологами и физиологами накоплен богатый материал о строении и функциях ретикулярной формации. Установлено, что структурные элементы ретикулярной формации локализуются в целом ряде мозговых образований, начиная с промежуточной зоны шейных сегментов спинного мозга (VII пластина), и заканчивая некоторыми структурами промежуточного мозга (интраламинарными ядрами, таламическим ретикулярным ядром). Ретикулярная формация состоит из значительного числа нервных клеток (в ней содержится почти 9/10 клеток всего ствола мозга). Общие черты строения ретикулярных структур — наличие особых ретикулярных нейронов и отличительный характер связей.
Рис.
Что такое ретикулярная формация
1. Нейрон ретикулярной формации. Сагиттальный разрез ствола мозга крысенка.
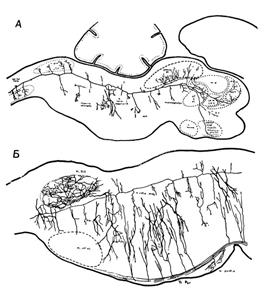
На рисунке А представлен только один нейрон ретикулярной формации. Видно, что аксон разделяется на каудальный и ростральный сегменты, большой протяженности, со множеством коллатералей. Б. Коллатерали. Сагиттальный разрез нижней части ствола мозга крысенка, показывающий соединения коллатералей большого нисходящего пути (пирамидный путь) с ретикулярными нейронами. Коллатерали восходящих путей (сенсорные пути), отсутствующие на рисунке, соединяются с ретикулярными нейронами подобным же образом (по Шейбэлу М. Э. и Шейбэлу А. Б.)
Наряду с многочисленными отдельно лежащими нейронами, различными но форме и величине, в ретикулярной формации головного мозга имеются ядра. Рассеянные нейроны ретикулярной формации прежде всего играют важную роль в обеспечении сегментарных рефлексов, замыкающихся на уровне ствола головного мозга. Они выступают в качестве вставочных нейронов при осуществлении таких рефлекторных актов, как моргание, роговичный рефлекс и т. д.
Выяснено значение многих ядер ретикулярной формации. Так, ядра, расположенные в продолговатом мозге, имеют связи с вегетативными ядрами блуждающего и языкоглоточного нервов, симпатическими ядрами спинного мозга, они участвуют в регуляции сердечной деятельности, дыхания, тонуса сосудов, секреции желез и т. д.
Установлена роль голубого пятна и ядер шва в регуляции сна и бодрствования. Голубое пятно, находится в верхнелатеральной части ромбовидной ямки. Нейроны этого ядра продуцируют биологически активное вещество — норадреналин, который оказывает активирующее воздействие на нейроны вышележащих отделов мозга. Особенно высока активность нейронов голубого пятна во время бодрствования, во время глубокого сна она угасает почти полностью. Ядра шва располагаются по срединной линии продолговатого мозга. Нейроны этих ядер вырабатывают серотонин, который вызывает процессы разлитого торможения и состояние сна.
Ядра Кахаля и Даркшевича, относящиеся к ретикулярной формации среднего мозга, имеют связи, с ядрами III, IV, VI, VIII и XI пар черепных нервов. Они координируют работу этих нервных центров, что очень важно для обеспечения сочетанного поворота головы и глаз. Ретикулярная формация ствола головного мозга имеет важное значение в поддержании тонуса скелетной мускулатуры, посылая тонические импульсы на мотонейроны двигательных ядер черепных нервов и двигательных ядер передних рогов спинного мозга. В процессе эволюции из ретикулярной формации выделились такие самостоятельные образования, как красное ядро, черное вещество.
По структурно-функциональным критериям ретикулярная формация делится на 3 зоны:
1. Медианную, расположенную по средней линии;
2. Медиальную, занимающую медиальные отделы ствола;
3. Латеральную, нейроны которой лежат вблизи сенсорных образований.
Медианная зона представлена элементами шва, состоящие из ядер, нейроны которых синтезируют медиатор – серотонин. Система ядер шва принимает участие в организации агрессивного и полового поведения, в регуляции сна.
Медиальная (осевая) зона состоит из мелких нейронов, которые не ветвятся. В зоне располагается большое количество ядер. Встречаются также крупные мультиполярные нейроны с большим числом густо ветвящихся дендритов. Они образуют восходящие нервные волокна в кору больших полушарий и нисходящие нервные волокна в спинной мозг. Восходящие пути связи медиальной зоны оказывают активирующее влияние (прямо или опосредованно через таламус) на новую кору. Нисходящие пути оказывают тормозящее влияние.
Латеральная зона – к ней относятся ретикулярные образования, расположенные в стволе мозга вблизи сенсорных систем, а также ретикулярные нейроны, лежащие внутри сенсорных образований. Основным компонентом этой зоны является группы ядер, которые примыкают к ядру тройничного нерва. Все ядра латеральной зоны (за исключением ретикулярного латерального ядра продолговатого мозга) состоят из нейронов малой и средней величины и лишены крупных элементов. В этой зоне располагаются восходящие и нисходящие пути, обеспечивающие связь сенсорных образований с медиальной зоной ретикулярной формации и моторными ядрами ствола. Эта часть ретикулярной формации является более молодым и возможно прогрессивнее, с ее развитием связан факт уменьшения объема осевой ретикулярной формации в ходе эволюционного развития. Таким образом, латеральная зона – это совокупность элементарных интегративных единиц, сформированных вблизи и внутри специфических сенсорных систем.
Рис. 2. Ядра ретикулярной формации (РФ) (по: Niuwenhuys еt. аl, 1978).
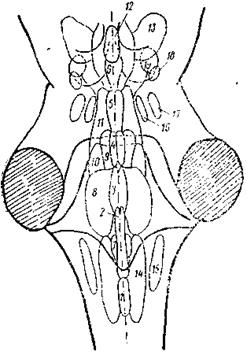 |
1—6 — медианная зона РФ: 1—4— ядра шва (1 — бледное, 2 — темное, 3 — большое, 4— мостовое), 5 — верхнее центральное, 6 — дорсальное ядро шва, 7—13 — медиальная зона РФ: 7 — ретикулярное парамедианное, 8 — гигантоклеточное, 9 — ретикулярное ядро покрышки моста, 10, 11 — каудальное (10) и оральное (11) ядра моста, 12 — дорсальное покрышечное ядро (Гуддена), 13 — клиновидное ядро, 14— I5—латеральная зона РФ: 14 — центральное ретикулярное ядро продолговатого мозга, 15 — латеральное ретикулярное ядро, 16, 17 — медиальное (16) и латеральное (17) парабрахиальные ядра, 18, 19 — компактная (18) и рассеянная (19) части педункуло-понтийного ядра.
Благодаря нисходящим влияниям ретикулярная формация оказывает тоническое влияние и на мотонейроны спинного мозга, что в свою очередь повышает тонус скелетной мускулатуры, совершенствует систему обратной афферентной связи. В результате любой двигательный акт совершается значительно эффективнее, осуществляет более точный контроль за движением, но чрезмерное возбуждение клеток ретикулярной формации может привести к дрожанию мышц.
В ядрах ретикулярной формации находятся центры сна и бодрствования, и стимуляция тех или иных центров приводит или к наступлению сна, или к пробуждению. На этом основано применение снотворных. В ретикулярной формации расположены нейроны, реагирующие на болевые раздражения, идущие от мышц или внутренних органов. В ней также расположены специальные нейроны, которые обеспечивают быструю реакцию на внезапные, неопределенные сигналы.
Ретикулярная формация тесно связана с корой больших полушарий, благодаря этому формируется функциональная связь между внешними отделами ЦНС и стволом головного мозга. Ретикулярная формация играет важную роль как в интеграции сенсорной информации, так и в контроле над деятельностью всех эффекторных нейронов (моторных и вегетативных). Она имеет также первостепенное значение для активации коры больших полушарий, для поддержания сознания.
Необходимо отметить, что кора полушарий большого мозга, и в свою очередь, посылает по корково-ретикулярным путям импульсы в ретикулярную формацию. Эти импульсы возникают в основном в коре лобной доли и проходят в составе пирамидных путей. Корково-ретикулярные связи оказывают либо тормозное, либо возбуждающее действие на ретикулярную формацию ствола головного мозга, они осуществляют корректировку прохождения импульсов по эфферентным путям (отбор эфферентной информации).
Таким образом, между ретикулярной формацией и корой полушарий большого мозга имеется двусторонняя связь, которая обеспечивает саморегуляцию в деятельности нервной системы. От функционального состояния ретикулярной формации зависит тонус мускулатуры, работа внутренних органов, настроение, концентрация внимания, память и т. д. В целом ретикулярная формация создает и поддерживает условия для осуществления сложной рефлекторной деятельности с участием коры полушарий большого мозга.
Дата добавления: 2017-03-12; просмотров: 1472;
Похожие статьи:
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Билет 15
1. Формы (фрагменты) афферентного синтеза: Доминирующая мотивация; Обстановочная афферентация; Пусковая афферентация. Роль ретикулярной формации.
2. Быстрые и медленные мышечные волокна.
Вопрос 1
АФФЕРЕНТНЫЙ СИНТЕЗ — (соединение, составление) — процесс сопоставления, отбора и синтеза многочисленных и разных по функциональному значению афферентаций, вызванных различными воздействиями на организм, происходящий в ц. н. с., на основе к-рого формируется цель действия.
А. с. согласно теории функциональной системы Анохина — первая, универсальная, стадия любого целенаправленного поведенческого акта (см. Функциональные системы).
А. с. включает обработку 4 основных видов афферентных возбуждений.
1. Мотивационное возбуждение отражает доминирующую потребность организма, к-рая возникает под влиянием метаболических, гормональных, а у человека — и социальных факторов Мотивация играет решающую роль в формировании цели действия . Специфически повышая реактивность корковых нейронов с помощью ориентировочно-исследовательской реакции, мотивационное возбуждение способствует обработке и активному отбору сенсорной информации, необходимой для построения целенаправленного поведения.
2. Обстановочная афферентация представляет собой воздействие на организм всей совокупности внешних факторов, составляющих конкретную обстановку, на фоне к-рой развертывается приспособительная деятельность. Обстановочная афферентация формируется не только постоянными компонентами обстановки, но и рядом последовательных афферентных воздействий на организм. Характерная особенность обстановочной афферентации состоит в том, что она придает специфику будущей поведенческой реакции, обеспечивая ее приспособительное значение только в данной обстановке.
Наиболее отчетливо роль обстановочной афферентации проявляется в опытах с условными рефлексами. В этих случаях на один и тот же условный раздражитель животное отвечает условной оборонительной реакцией в одной экспериментальной камере и условной пищевой — в другой (или в одной и той же экспериментальной камере утром животное отвечает пищевой реакцией, а вечером — оборонительной).
На стадии афферентного синтеза решаются вопросы “что делать?”, “как делать?”, “когда делать?”.
Пусковая афферентация
Представляет собой специальный раздражитель, собственно запускающий поведенческую реакцию. Значение пускового раздражителя состоит в том, что он призван обозначить момент начала поведенческой реакции.
Целенаправленное поведение может начинаться и без явного пускового стимула. Примерами таких реакций являются регулярно совершающиеся физиологические отправления (еда, сон, дефекация, мочеиспускание и др.), приуроченные к определенным периодам суток.
Афферентный синтез осуществляется на основе следующих нейрофизиологических механизмов:
1) механизмы восходящих активирующих влияний подкорковых образований на кору головного мозга. Это прежде всего, активирующие влияния гипоталамуса к лобным отделам коры, через передние ядра таламуса, что отражает мотивационные возбуждения. Аналогичным образом воздействуют другие лимбические системы. Вторыми по активирующему значению являются ретикулярные структуры среднего мозга и моста, которые обеспечивают соответствующий уровень бодрствования.
2) механизмы конвергенции возбуждений различного качества на нейронах коры и подкорковых структурах мозга. В частности, мультисенсорная конвергенция от поверхностей (зрительная, тактильная, слуховая, температурная и др.); мультибиологическая конвергенция, связанная с определенными состояниями (голод, боль и т.п.) и др.;
3) интеграцию мотивационных, обстановочных и пусковых афферентаций на нейронах коры мозга;
4) механизмы формирования доминанты, за счет которых подавляется текущая деятельность и удерживается вновь сформированная поведенческая реакция.
Роль ретикулярной формации
Ретикулярная формация характеризуется относительно малой возбудимостью. Эффекты ее раздражения появляются через большой латентный период, она медленно реагирует и остается активной в течение продолжительного времени после прекращения раздражения (длительное последействие). Ретикулярная формация облегчает или подавляет фазные движения и напряжение скелетных мышц, вызываемые мотонейронами спинного мозга, а также движения, вызванные с коры больших полушарий . Ретикулярная формация среднего и промежуточного мозга облегчает рефлекторные движения животных, раздражение промежуточного мозга тормозит двигательные рефлексы спинного мозга.
Боковые отделы ретикулярной формации варолиева моста и среднего мозга облегчают, а средние ее отделы в продолговатом мозге тормозят двигательные рефлексы. Облегчение и торможение зависят также от интенсивности и продолжительности раздражения ретикулярной формации. По гамма-нейронам она регулирует функции мышечных веретен, следовательно, обратную информацию из скелетных мышц. Она изменяет также возбудимость восходящих афферентных путей спинного мозга, что может снизить или прекратить постсинаптическое торможение. Тонические влияния ретикулярной формации вызывают ВПСП или ТПСП в мотонейронах спинного мозга. Она изменяет также передачу импульсов в мозговом стволе и одновременно с влиянием на скелетную мускулатуру вызывает сосудодвигательные, дыхательные, зрачковые и другие реакции.
Ретикулярная формация оказывает адаптационно-трофическое влияние на кору больших полушарий, подкорковые образования промежуточного мозга, мозжечок и спинной мозг. Существуют взаимные влияния этих отделов нервной системы, как возбуждающие, так и тормозящие. Она участвует в физиологических процессах сна и пробуждения, а также в эмоциях, в реакции напряжения («стресс») и др. Раздражение ретикулярной формации вызывает пробуждение спящих животных, а ее разрушение и выключение — глубокий сон у бодрствующих животных. Изучены взаимные влияния ретикулярной формации и коры больших полушарий .Установлено участие ретикулярной формации в образовании и протекании условных рефлексов
По симпатическим волокнам ретикулярная формация регулирует возбудимость и работоспособность скелетной мускулатуры, функциональное состояние нервной системы и органов чувств, оказывая на них адаптационно-трофическое влияние. Регуляция рефлексов позы и двигательных рефлексов, перемещающих тело, осуществляется по эфферентным гамма-волокнам иннервирующим проприоцепторы.
Ретикулярная формация регулирует вегетативные функции, деятельность внутренних органов. Она влияет на образование гормонов в гипофизе и других железах внутренней секреции и в ней концентрируются гормоны и медиаторы.
Афферентные волокна поступают в нее по симпатическим и блуждающим нервам.
Частьклеток ретикулярной формации среднего мозга и варолнева моста возбуждается адреналином и норадреналином (адренореактивные системы) а другая часть, расположенная в промежуточном мозге, несколько выше среднего мозга, возбуждается ацетилхолином и его производными (холинореактивные системы). Адренореактивные системы среднего мозга и варолиева моста облегчают наступление двигательных рефлексов, а адренореактивные системы продолговатого мозга тормозят спинномозговые рефлексы. Адреналин возбуждает и холинореактивные системы. Предполагается, что действие ацетилхолина и его производных менее ограничено, чем действие адреналина, и охватывает многие области головного мозга. Действие ацетилхолина на ретикулярную формацию противоположно его периферическому влиянию на внутренние органы. Ретикулярную формацию среднего и продолговатого мозга возбуждает углекислота.
Гормоны и медиаторы действуют на функцию больших полушарий как непосредственно, так и посредством ретикулярной формации. Таким образом, ретикулярная формация мозгового ствола — подкорковый центр вегетативной нервной системы.
Вопрос 2.
Читайте также:
Анатомия ретикулярной формации
Ретикулярная формация(formatio reticularis; лат. reticulum сетка; синоним ретикулярная субстанция) — комплекс клеточных и ядерных образований, занимающих центральное положение в стволе головного мозга и в верхнем отделе спинного мозга. Большое количество нервных волокон, пронизывающих ретикулярную формацию в различных направлениях, придает ей вид своеобразной сеточки, что послужило основанием для названия этой структуры.
Вся ретикулярная формация может быть подразделена на каудальный, или мезэнцефалический, и ростральный, или таламический, отделы. Каудальный отдел ретикулярной формации определяет диффузную, неспецифическую систему влияний на сравнительно обширные отделы и зоны головного мозга, тогда как ростральный отдел ретикулярной формации — специфическую систему, оказывающую относительно локальные влияния на определенные зоны коры больших полушарий. Диффузность (или специфичность) ретикулярной формации проявляется и в характере модальности нервно-импульсных влияний. Таким образом ретикулярная формация — это универсальная система, обусловливающая функциональное состояние всех отделов головного мозга и влияющая на все виды нервной деятельности, т.е. ее можно представить как «мозг в мозге».
Несмотря на многообразие форм влияния ретикулярной формации на деятельность нервной системы, можно выделить два основных направления воздействия ретикулярной формации: ретикулоспинальные влияния и ретикулокортикальные взаимоотношения.
Ретикулоспинальные влияния носят облегчающий или тормозной характер и играют важную роль в координации простых и сложных движений, в реализации влияний психической сферы на осуществление сложной двигательной поведенческой деятельности человека.
Ретикулокортикальные взаимоотношения разноплановы. Из клинической практики известно, что при поражении определенных отделов ствола головного мозга наблюдаются снижение двигательной активности, сонливость, ареактивность, нарушение смены состояний сна и бодрствования, подавление психической деятельности, т.е. снижение активирующих влияний на процессы корковой интеграции. Показано также, что раздражение определенных участков ретикулярной формации вызывает в обширных зонах коры больших полушарий реакцию активации. Эти данные позволили сформулировать концепцию о диффузной, восходящей, активирующей системе ретикулярной формации.
Анатомия среднего мозга
Сре́дний мозг (лат. Mesencephalon) — отдел головного мозга, древний зрительный центр. Включен в ствол головного мозга.
Мозг человека
1. Cerebrum — большие полушария
2. Mesencephalon — средний мозг
3. Pons — мост
4. Medulla oblongata — продолговатый мозг
5. Cerebellum — мозжечок
Вентральную часть составляют массивные ножки мозга, основную часть которых занимают пирамидные пути. Между ножками находится межножковая ямка (лат. fossa interpeduncularis), из которой выходит III (глазодвигательный) нерв. В глубине межножковой ямки — заднее продырявленное вещество (лат. substantia perforata posterior).
Дорсальная часть — пластинка четверохолмия, две пары холмиков, верхние и нижние (Шаблон:Culliculi superiores & inferiores).
Ретикулярная формация мозга: строение и функции
Верхние, или зрительные холмики несколько крупнее нижних (слуховых). Холмики связаны со структурами промежуточного мозга — коленчатыми телами, верхние — с латеральными, нижние — с медиальными. С дорсальной стороны на границе с мостом отходит IV (блоковый) нерв, сразу же огибает ножки мозга, выходя на переднюю сторону. Четкой анатомической границы с промежуточным мозгом нет, за ростральную границу принята задняя комиссура.
Внутри нижних холмиков находятся слуховые ядра, туда идет латеральная петля. Вокруг сильвиева водопровода — центральное серое вещество (лат. substantia grisea centralis).
В глубине покрышки среднего мозга (под четверохолмием) находятся ядра глазодвигательных нервов, красные ядра (лат. nuclei rubri, управление движением), черное вещество (лат. substantia nigra, инициация движений), ретикулярная формация.
Нервные центры продолговатого мозга
В продолговатом мозге находятся центры:
— дыхательный и сосудодвигательный;
— возбуждающий слезоотделение;
— регулирующие сердечную деятельность;
— тормозящие деятельность сердца;
— вызывающие выделение желчи и сокращение желудочно-кишечного тракта;
— секрецию слюнных, поджелудочных и желудочных желез.
©2015-2018 poisk-ru.ru
Все права принадлежать их авторам. Данный сайт не претендует на авторства, а предоставляет бесплатное использование.
Нарушение авторских прав и Нарушение персональных данных
Больше информации по теме
 Достоверность доказательств в арбитражном процессе
Достоверность доказательств в арбитражном процессе Кабальный холоп
Кабальный холоп Княжеская власть в древнерусском государстве
Княжеская власть в древнерусском государстве Структура защитительной речи
Структура защитительной речи Классификация фактов
Классификация фактов Виды социального обеспечения военнослужащих
Виды социального обеспечения военнослужащих Виды режимов рабочего времени
Виды режимов рабочего времени Возникновение франкского государства
Возникновение франкского государства Полномочия суда апелляционной инстанции в арбитражном процессе
Полномочия суда апелляционной инстанции в арбитражном процессе